Outline
The problem of free will lies at the heart of modern scientific studies of
consciousness. An influential series of experiments by Libet has suggested that
conscious intentions arise as a result of brain activity. This contrasts with traditional concepts of free will, in which the mind controls the body. A more recent
study by Haggard and Eimer has further examined the relation between intention
and brain processes, concluding that conscious awareness of intention is linked
to the choice or selection of a specific action, and not to the earliest initiation of
action processes. The exchange of views in this post further explores the relation between conscious intention and brain activity
The recordable cerebral activity (readiness-potential, RP) that precedes a freely voluntary, fully endogenous motor act was directly compared with the reportable time (W) for appearance of the subjective experience of 'wanting' or intending to act. The onset of cerebral activity clearly preceded by at least several hundred milliseconds the reported time of conscious intention to act. This relationship held even for those series (with 'type II' RPs) in which subjects reported that all of the 40 self-initiated movements in the series appeared 'spontaneously' and capriciously.
Data were obtained in at least 6 different experimental sessions with each of 5 subjects. In series with type II RPs, onset of the main negative shift in each RP preceded the corresponding mean W value by an average of about 350 ms, and by a minimum of about 150 ms. In series with type I RPs, in which an experience of preplanning occurred in some of the 40 self-initiated acts, onset of RP preceded W by an average of about 800 ms (or by 500 ms, taking onset of RP at 90 per cent of its area). Reports of W time depended upon the subject's recall of the spatial 'clock-position' of a revolving spot at the time of his initial awareness of wanting or intending to move.
Two different modes of recall produced similar values. Subjects distinguished awareness of wanting to move (W) from awareness of actually moving (M). W times were consistently and substantially negative to, in advance of, mean times reported for M and also those for S, the sensation elicited by a task-related skin stimulus delivered at irregular times that were unknown to the subject. It is concluded that cerebral initiation of a spontaneous, freely voluntary act can begin unconsciously, that is, before there is any (at least recallable) subjective awareness that a 'decision' to act has already been initiated cerebrally. This introduces certain constraints on the potentiality for conscious initiation and control of voluntary acts.
Voluntary action is fundamental to human existence. Most of us navigate through
our daily lives with the belief that we have conscious free will: that is, we have
conscious intentions to perform specific acts, and those intentions can drive our
bodily actions, thus producing a desired change in the external world. The doctrine of conscious free will seems, at first sight, to be strongly dualist, and therefore incompatible with the reductionism of modern brain science: how can a
mental state (my conscious intention) initiate the neural events in the motor areas
of the brain that lead to my body movement? Modern neuroscience would reverse
these causal roles, and would describe conscious intention as a consequence or
correlate of neural preparation of action.
Despite these scientific worries, the concept of conscious free will remains deeply rooted in our individual lives and in
our societies. As Hume (1739/1955) observed:
On this is founded our belief in witnesses, our credit in history, and indeed in all
kinds of moral evidence, and almost the whole conduct of life (p. 182).
Perhaps the most interesting scientific attempt to break out of this impasse comes
from the experiment of Libet et al. (1983). Before describing the experiment in
detail, I [PH] would like to single out two reasons why I believe it has been so
important. Libet was the first to attempt a scientific psychophysiology of free
will. His experiment brought scientific method to a question that had previously
been purely philosophical.
Using this method, he produced data that support the
traditional neuroscience view, and deeply undermine the concept of conscious
free will: preparatory brain activity causes our conscious intentions. The second
outstanding feature of Libet's study is the insight that we may have a conscious
veto over the acts our brain has previously unconsciously prepared (so called
'free won't'), even if we lack conscious free will. This brilliant revision of the traditional concept of free will saves most of its desirable corollaries, such as individual liberty and moral responsibility, while maintaining compatibility with
modern neuroscience.
I will next briefly summarise the experiment which
brought Libet to these conclusions, and will then introduce the controversy over
selection
Libet and colleagues asked subjects to fixate a small clock hand, which rotated
once every 2.56 s. Subjects then made a voluntary movement (they were
instructed to flex their wrist 'freely and capriciously') at a time which they themselves chose. The clock continued to rotate for a random interval after the voluntary movement, and then stopped. Subjects then reported the position of the clock
at which they first became aware of the will to move. Libet termed this the 'W
judgement', and took it as the first moment of conscious intention. In other conditions, subjects judged the time at which the actual movement began ('M judgement'): for the present discussion the W judgement is the most important, though
we shall return to the M judgement later.
The exact moment at which the action began was calculated by measuring the
electrical activity in the muscles involved. The preparatory activity in the motor
areas of the brain (the readiness potential - RP) was also calculated by measuring electrical activity with a scalp electrode placed over the motor cortex, and
averaging epochs of data prior to each voluntary action. The RP is a wellestablished gradual increase in electrical activity in the motor cortical regions,
which characteristically precedes willed actions by 1 s or more, and is strongly
related to the effort, thought and attention required to generate the action.
The combination of psychophysical estimates and physiological recording
gave Libet and colleagues the measures required to address the question of conscious free will. Specifically, the temporal order of the two measures allowed
Libet and colleagues to investigate which event was cause and which was the
effect. If the moment of conscious intention preceded the onset of the readiness
potential, then the concept of conscious free will would be tenable: the early conscious mental state could initiate the subsequent neural preparation of movement.
But if the moment of conscious intention followed the onset of the readiness
potential, then conscious free will cannot exist: a conscious mental state must be a
consequence of brain activity, rather than the cause of it.
Libet et al.'s data showed the latter pattern. Their results can be stated quite
simply, and are shown schematically in Figure 1. The readiness potential began
around 1000-500 ms before the onset of actual body movement.
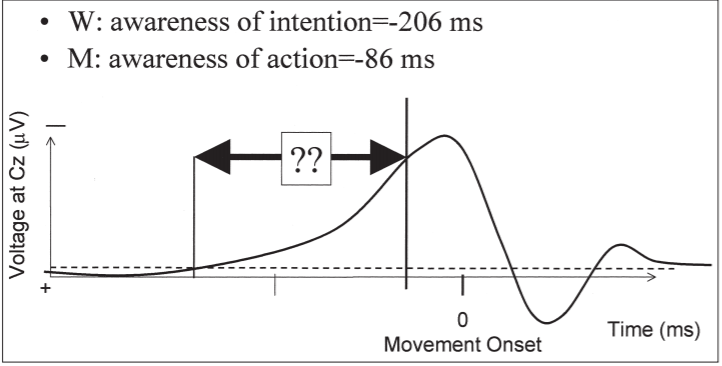
Figure 1. Schematic showing the important features of the results of Libet et al. (1983)
The exact time of onset differed between two different types of RP observed in the data, which
Libet called RPI and RPII. Type I RPs had earlier onsets than type II RPs. Type I
RPs were found in blocks where subjects reported the experience of planning and
consciously preparing their actions, on at least some trials. Type II RPs were
found in blocks where subjects reported that their actions were unplanned, and
that they occurred more spontaneously.
The key result for the present purpose, however, is the interval between RP
onset and W judgement. Subjects' W judgements showed that they only experienced conscious intention an average 206 ms before movement onset. This is
around 350 ms after the onset of type II RPs, and around 500 ms after the onset of
type I RPs.
Thus, the brain is preparing the purportedly 'free' action significantly
before the subject himself is aware they he intends to move. This temporal gap
poses a difficulty for the traditional concept of free will.
Several critiques of the above experiment were published in 1985, in response
to a target article in Behavioral and Brain Sciences (Libet, 1985). These critiques
raise a number of problems with Libet's method, but only one of these really concerns us here. People are generally poor at judging the synchrony of two events
occurring in different perceptual modalities (Sternberg and Knoll, 1973) or perceptual streams. In particular, events in an attended stream appear to occur earlier
than simultaneous events in an unattended stream (the so-called Prior Entry phenomenon).
Since Libet's subjects presumably divided their attention in varying
proportions between the external clock and their own internal states in order to
make the W judgement, the precise numerical value of 206 ms must be treated
with caution. In my own view, criticisms of the Libet method based on attentional
biases have been overstated. Estimates of the prior entry effect run from 70 ms
(Sternberg and Knoll, 1973) down to 12 ms (Shore et al., 2001; Haggard and
Johnson, 2001). Even the largest of these values is an order of magnitude smaller
than Libet's gap between readiness potential and W judgement.
Specificity and Intentions
Our own point of entry to the debate was twofold (Haggard and Eimer, 1999).
Firstly, we wanted to investigate the causal relations between brain preparation
and conscious awareness in more detail. Second, we wanted to link the two types
of event by an inference that did not depend on the absolute numerical value of a
timing judgement (which henceforth I shall call a 'Libet estimate'). Our experiment included some other factors, such as a comparison between fixed-choice
and free-choice action, which are not of interest here.
The original 1983 paper suggested that the readiness potential was the cause of
subsequent conscious awareness: as the subtitle put it: 'The unconscious initation
of a freely voluntary act'. The basis for this causal argument was the temporal
precedence of readiness potential (RP) onset over conscious intention, as
described above. Temporal precedence may be important for the controversy
between mind-to-brain vs brain-to-mind causation, but it is a necessary rather
than a sufficient condition for a causal relation. In particular, there might well be
other neural premotor events that are more plausible than the RP as causes of conscious intention. We particularly wished to investigate which of the readiness
potential or the lateralised readiness potential (LRP) is a more plausible cause of
conscious intention.
Therefore, we now briefly explain the meaning and importance of the LRP.
The early portions of the RP are symmetrical over both hemispheres, but the
later portions of the RP show an increasing shift towards the hemisphere
contralateral to the hand that will make the forthcoming action, typically beginning some 500 ms prior to movement onset. This Lateralised Readiness Potential
(LRP) can be calculated by subtraction of electrode signals located symmetrically
over the two motor areas (Eimer, 1998). The LRP has a particular psychological
significance in situations where the subject must choose between a left- and a
right-handed action: once the LRP has begun, the selection of which action to
make must be complete. That is, by LRP onset the intention has progressed from
abstract stage ('Do something or other!') to drive a specific movement ('Do precisely this!').
Comparing the plausibility of two candidate causes often comes down to comparing how well each candidate correlates with the putative effect (Mill's method
of concomitant variation).
We relied on the fact that these experiments always
involve collecting large numbers of repeated trials. The variability across trials in
both W judgements and RPs is typically high. Therefore, we related both RP and
LRP onsets to W judgements. For example, if the RP is truly the cause of conscious intentions, then trials which happen to have early RP onsets (i.e., onsets
occurring long before the movement itself) should also have early conscious
intentions. Ideally, this would be done by predicting W judgement value from the
RP or LRP onset value in each trial. However, the RP and LRP measures have a
poor signal-to-noise ratio, and can only be obtained by averaging across several
trials. Therefore, we were obliged to apply Mill's logic backwards: we classified
each subject's trials according to whether the W judgement was early or late
relative to that subject's median W value, and calculated RPs and LRPs corresponding to these early and late judgements respectively.
Briefly, we found RP onset did not covary with W judgements: trials with early
W judgements in fact showed later RP onsets than trials with late judgements.
LRP onset, however, did covary with W judgement: trials with earlier W judgements had earlier LRP onsets than trials with later W judgements. The pattern of
results is shown in Table 1. From these data we concluded that the RP could not be
the cause of conscious intention, but the LRP could be a possible cause of conscious intention. Since the motor system must have selected which specific movement to perform by the time that the readiness potential lateralises, we concluded
that conscious intentions were related to specific rather than general preparation
for action.
|
|
Early W Trials
|
Early W Trials
|
|
Mean W
|
-530
|
-179
|
|
LRP onset
|
-906
|
-713
|
Implications
The purpose of this post is to discuss the implications of both the original result,
and of our re-enactment for the cognitive neuroscience of willed action. Libet's
original result suggested that conscious intentions were a consequence of the very
early general preparation for movement by the brain. Our result, in contrast,
suggests that conscious awareness of intention should be attributed only at a later
stage in the genesis of an action: namely after the selection of a specific motor
implementation of the action by a left or right handed keypress.
The question of what causes conscious intentions is important for several reasons: hence this discussion.
First, we need to know what brain event, if any,
causes conscious intentions, in order to shed light on the traditional problem of
conscious free will. Second, identifying the neural basis of intentions may prove
useful in understanding disorders of intentional action such as those which occur
in schizophrenic patients with hallucination and delusions.
In this section, I will discuss several implications of the specificity result of
Haggard and Eimer (1999) described above. I believe the result has implications
for the evolutionary value of action awareness, for the traditional free will problem, and for the unity of conscious awareness. Finally, and more speculatively, I
think the specificity result may have implications for Libet's salvaging of free
will through the conscious veto. These will be discussed in turn.
1. Does conscious awareness arise at the stage of specification?
Why might conscious awareness arise at the stage where the CNS selects the specific movement which will be used to achieve the action? Computational
approaches to motor control have generally agreed that movement selection is the
most informationally difficult problem in action. In most everyday actions, there
are several different ways to achieve a desired goal. For example, I could reach
for the milk bottle with my left or my right hand, I could take a couple of steps to
get closer to the fridge, or I could extend my arm to the limit of its reach: in each
case I succeed in the action of getting the bottle, though I do so using very different movements.
The CNS faces the problem of choosing just one of the infinite
set of possible movements. This problem is ill-posed: there is no unique solution.
This is known in psychology as the problem of motor equivalence (Hebb, 1949),
and as kinematic redundancy in the specific case of computational motor control
(Wolpert and Ghahramani, 2000). The limited action repertoire of so-called
'intelligent' robots suggests that the human motor system uses quite sophisticated
information-processing to solve the problem. Our specificity result suggests that
conscious awareness may be a consequence of this information-processing.
Awareness of what we will do is tied to the selection of specifically how we will
do it. Other accounts of consciousness, based on very different domains, have
suggested that conscious awareness arises when particular kinds of computational processes occur (Jack and Shallice, in press). Our specificity result similarly suggests that consciousness of intention may be related to a particular kind
of processing involved in movement selection.
This view makes clear predictions which could be tested in subsequent
research. The phenomenology of intention should be strongest when the movement selection computations are most intensive. For example, awareness of
intention may be more vivid, and may possibly occur earlier for actions where the
subject must carefully select between a large number of alternative movements,
for example when reaching for one object among a number of distractors.
2. Specificity and the free will problem
Our result used a correlational approach, based on Mills' method of concomitant
variation, to identify the cause of conscious awareness of intention. Famously,
however, correlation is not causation. In particular, a correlation between two
events does not identify which is the cause and which is the effect. In our result,
the only grounds for inferring that the LRP causes awareness of intention, rather
than the other way around, is the temporal precedence argument originally used
by Libet: causes precede their effects. Temporal precedence is particularly germane here, because, as described above, the numerical values of Libet estimates
are not a robust guide to the exact times of mental events.
Biases in cross-modal synchronisation, prior entry, and individual differences in time perception could
all make Libet estimates poor guides to the actual time at which subjects experience an awareness of
when they will act (for the sake of this discussion, we will
grant the assumption that such a distinct mental event occurs in the first place).
Anyone wishing to rescue the traditional concept of conscious free will should
pay close attention to these methodological issues. Libet's basis for rejecting the
traditional concept of free will as the initiator of action was the long gap between
RP onset and W awareness (see Figure 1). It is this gap, or temporal precedence,
that justifies a brain-mind rather than a mind-brain direction of causation. We
have now suggested that LRP, rather than RP onset is the physiological event of
interest, and LRP onset of course occurs much later than RP onset. Therefore, the
gap between brain and mind events that needs to be explained away by the proponent of conscious free will is much narrower judging from our result than Libet
originally reported.
Inspection of Table 1 shows that if numerical W estimates can
be shown to be delayed by, say 500 ms, relative to the actual conscious experience
of intention, then the temporal precedence of LRP over W awareness would be
reversed. The free will theorist could suggest that conscious intentions cause the
brain processes of movement selection (of course, the other arguments against
such a view would not thereby be answered).
I doubt that quirks of human estimation, such as the prior entry effect, could by
themselves produce errors of as much as 500 ms in estimation. But there may be
other reasons why reports of conscious experience are delayed relative to the
actual experience itself. For example, the P-centre phenomenon originally discovered for speech stimuli (Morton et al., 1976) may apply equally to internal
events. The P-centre phenomenon refers to the fact that the perceived onset of a
speech stimulus lags its physical onset, and seems to be attracted by the 'centre'
of the stimulus.
Intentions might likewise be extended in time. The requirement to
judge their onset as a discrete event is clearly somewhat arbitrary and difficult:
people may resort to judging the centre of some extended process. The earliest
stages of that process could perhaps precede LRP onset.
Again, it is useful to think of empirical approaches to the question of temporal
precedence. An experiment identifying a neural event occurring at the precise
moment of conscious awareness could be particularly important.
For example,
locking EEG records with respect to W judgements on each trial, as opposed to
locking them to movement onset, might reveal a particular neural event, let us call
it Wn, which occurs close to the time of W. If Wn were found to follow both RP and
LRP onset, and also to covary in time with them, then I believe this would be
strong evidence for brain-mind causation, rather than the mind-brain causation
hypothesised by conscious free will accounts
A second possibility for identifying the precise temporal relations between
brain activity and awareness of intention is suggested by a fascinating but
little-known study performed by Fried et al. (1991).
Those authors stimulated the frontal cortex intracranially, through surface electrodes implanted
as part of a surgical procedure for intractable epilepsy. Stimulation at some of their more anterior
electrode sites, corresponding to Brodmann's area 6, caused their patient's to
report an urge to move a specific body part, or a feeling that they were about to
move. Weak stimulation typically evoked such reports of conscious intention,
while stronger stimulation evoked actual movements of the same body part.
The similarity of these reports to the W judgement is interesting, though of course
Fried's subjects' reports were not reports of the time of awareness. Moreover,
brain processes induced by experimental stimulation and the spontaneous brain
processes that are measured with EEG may be quite different. Nevertheless, it is
tempting to think that Fried's patients experienced passively what Libet's and our
subjects achieved themselves. Fried's result is clearly consistent with a brain-
mind direction of causation: conscious awareness of intention is the consequence
of frontal brain activity.
Unfortunately, the Fried et al. study was not chronometric: we have no clear
idea of the interval between electrical stimulation and the subject first feeling an
urge to move. A replication of the Fried study, in which subjects made rapid
choice responses to their own awareness of intention would be very interesting
scientifically. It would also provide timing estimates which did not rely on the
Libet clock, and would therefore avoid the problems of cross-modal synchronisation, prior entry, etc
Specificity and the relation between Libet's W and M
A useful framework for thinking about voluntary action and conscious awareness
is given in Figure 2. The left hand side shows the brain events occurring with voluntary action, while the right hand side shows the conscious experiences that may
be associated with voluntary action. I wish to make the strong claim that the relations between the neural events and conscious events are not one-to-one, but are
many-to-many. Libet et al.'s (1983) paper was the first to attempt a
psychophysiology of free will.
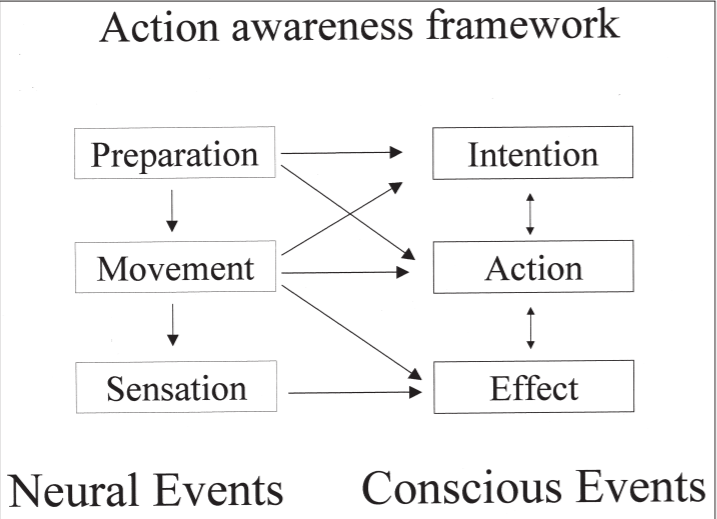
Figure 2. A framework for understanding the conscious awareness of action. See text for details.
It made a link between neural preparation and
awareness of intention. Our specificity result suggests that awareness of intention
depends also on the actual movement made, giving a many-to-one mapping from
neural events to conscious events.
Other studies (Haggard and Magno, 1999)
have suggested that judgements of when we actually move (Libet's 'M judgement') depend on the plan or intention to move, as well as on actual muscular
contraction. This implies a one-to-many mapping, whereby neural preparation
contributes to awareness of both intention and action. In more recent work (Haggard et al., in press), we have reported a binding effect in operant action, whereby
the conjunction of intentional actions and their effects leads to these events being
perceived closer together in time than the same events presented singly. In this
sense, all the neural events on the left hand side of Figure 2 seem to contribute to
any conscious event studied. That is, conscious experiences surrounding action
are integrated from a series of actual events.
I believe this integrative operation of action awareness explains an interesting
feature of Libet's original data. In the 1983 paper, Libet focussed on explaining
the gap between RP onset and W. However, I believe his data reveals a second gap
which is equally intriguing for students of consciousness. In Libet's data, the
interval between the neural events of preparation (RP onset) and actual movement is probably around 1 s (Libet's criteria for detecting RP onset were rightly
conservative, so his reported measures are around 500-900 ms, depending on
whether an RP is type I or type II).
Interestingly, the equivalent interval between
the corresponding conscious events W and M, is much shorter (Figure 3). Thus,
an extended period of neural activity produces a much narrower, more discrete
conscious awareness. This again suggests an integrative operation of conscious
awareness, which we shall call intentional binding elsewhere (Haggard et al., inpress).
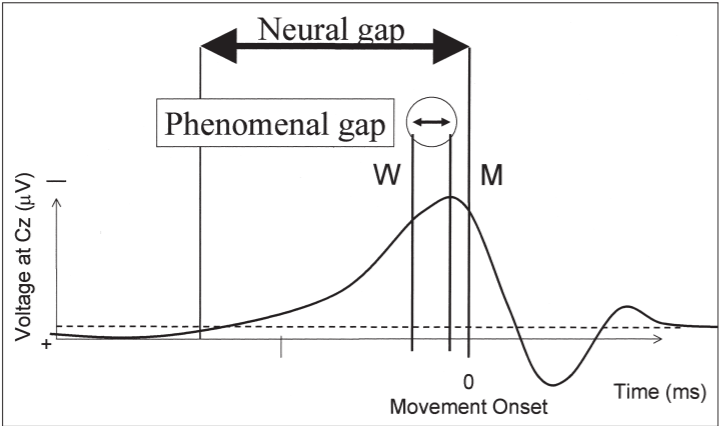
Figure 3. Conscious experience of the events occurring during voluntary action is compressed
in time relative to the actual occurrence of those events. Note that the neural gap between RP
onset and movement onset is experienced as a much shorter interval between W and M.
It seems likely that this integration has the function of producing unified
experience of our own voluntary action across space and time, though we must be
somewhat careful about this claim because Libet judgements do not test the unity
of experience directly.
If I am right about the integrative operation of action awareness,
then the specificity result also makes sense. Asking a subject to make M or W judgements does
not guarantee that they have a distinct underlying conscious experience
corresponding to those judgement categories. Rather, people may have conscious
awareness of a compressed core of events clustered around the action itself.
Movement specification lies closer to that core than the earliest onset of the neural activity.
4. Specificity and the conscious veto
Having disproved the traditional concept of free will, Libet salvaged some of its
important consequences such as attribution of actions to agents, free choice and
personal responsibility. He did this by noting that, while actions appear not to be
freely initiated, they may be freely stopped. He suggested that there was sufficient
time between W awareness and movement onset for a conscious veto to operate.
I suggest there may be a similarity between conscious veto and the relation
between W and movement specification.
In a choice situation like our experiment, W awareness seems to be related to modification of action. One reason for
tying W awareness to specification could be to allow an option for final, conscious decision on the question 'Is that really the right way to achieve what I
intend to do?' Libet's conscious veto has a similar but more radical role of asking
whether the action should be cancelled entirely. That is, Libet's veto corresponds
to the internal question 'Do I really want to realize this intention?' It seems to me
that the two questions should be related: once an intention has be translated to a
specific action plan, and has reached conscious awareness, a whole series of
checks and internal mental simulations should begin at many levels in the motor
system.
These checks would monitor both the desirability of the action and its
effect (Libet's veto), and also whether the specific action plan is the best way to
achieve the effect (Haggard's specificity). It is unclear which monitoring processes reach conscious awareness, and under what circumstances. The philosophical implications of this multiplicity of monitoring processes also remain to be
worked out.
Haggard advances the idea that conscious awareness of intention is linked to the choice of a specific action, rather than the initiation of an action process. Libet's experiments had suggested that readiness potentials for action happened before the consciousness of volition to act. The experiments appeared to confirm a pre-existing neuroscientific bias in favour of the idea that consciousness had little or no role to play.
His theory seemed to show that unconscious preparations for action preceded awareness of the wish to act. He did however try to save freewill by positing a conscious veto that could prevent the action being performed. This qualification of his initial finding proved a good deal less popular with the neuroscientific establsihment.
Haggard's first substantial criticism of the Libet experiment is that the fact that the readiness potential came before the conscious awareness is necessary but not sufficient to establish a causal relationship. Other neural events might also impact consciousness. Haggard distinguishes between the initial readiness potential and the lateral readiness potential (LRP), which comes when the subject has decided which hand to use in the Libet experiment.
The LRP was found to have a closer correlation with perceived moment of conscious volition than the original readiness potential (RP) highlighted by the Libet experiment. This suggested that the conscious intention was related to specific rather than a general preparation for action. Haggard asks why it is might be that consciousness was associated with the selection of a specific action. Computational work relative to motor control has established that movement selection is the most difficult problem in action, because there are an infinite number of possible movements that could achieve a particular end, and there is no unique solution.
Haggard suggests a relation between the necessary information processing for movement selection and conscious information processing. He suggests that his proposition could be tested by experiment, by matching experience of intention to movement selecting processing in the brain. In terms of the Libet veto, Haggard connects this to the decision as to whether a specific action selected at the point of the readiness potential is in fact the best possible action, or a suitable action at all.